
|
In Search of Ecological Relevancy: A Review of Landscape Fragmentation Metrics and Their Application for the Fundy Model Forest Submitted to the Group 1 (Biodiversity) Working Group
ABSTRACT
2.0 Basic Concepts: Biodiversity, Landscape Ecology, and Fragmentation 3.0 Landscape Metrics
3.2 Patch Size Metrics 3.3 Edge Metrics 3.4 Landscape Configuration Metrics 3.5 Patch Shape Metrics 3.6 Summary 5.0 The Relative Importance of Landscape Pattern to Selected Taxa 6.0 Conclusion and Recommendations: Application of Fragmentation Metrics in the Fundy Model Forest Literature Cited Table 1. Generalized predicted direction of change as a result of human-induced disturbance Table 2. Summary of studies reporting landscape effects List of Figures Figure 1. Edge effect and Core Area 1.0 Introduction "Degree of forest fragmentation or connectedness of forest ecosystem components" is one of the principal indicators of forest biodiversity in the Fundy Model Forest (FMF)(Etheridge et al. 1999). The Canadian Council of Forest Ministers also identified fragmentation as one of the important indicators to monitor (CCFM 1995). Further, guidelines developed for the protection of native biodiversity in the FMF recommend landscape level management strategies such as minimum patch size and connectivity (Woodley and Forbes 1997). However, scant information exists on the rate of habitat fragmentation in the FMF. Very little ecologically-focussed monitoring of landscape composition or pattern has been conducted. Woodley (1993) examined habitat fragmentation in an Intensive Study Area surrounding Fundy National Park. He used a range of methods to measure the impact of fragmentation on ecological integrity. However, in recent years a number of new methods have emerged for measuring habitat fragmentation (McGarigal and Marks 1995, Baskent 1999). The purpose of this project is four fold: (1) to review the concepts of biodiversity, landscape ecology and fragmentation, (2) to review metrics that have been proposed to measure fragmentation, (3) to summarize studies that have applied metrics to landscapes, and (4) to summarize landscape ecological literature to determine which metrics are the most ecologically relevant. The utility of these metrics will be explored in relation to the habitat requirements of a range of forest-dependent taxa. This paper will provide the necessary basis for the initiation of forest landscape monitoring in the Fundy Model Forest. 2.0 Basic Concepts: Biodiversity, Landscape Ecology, and Fragmentation Biodiversity is the variety of life and the ecological and evolutionary processes that support it (Noss and Cooperrider 1994, Woodley and Forbes 1998). Biodiversity is often characterized at four scales: (1) genetic diversity, (2) species diversity, (3) community diversity, and (4) landscape diversity. These scales are interconnected because the processes that create biodiversity are multiple scale events (spatially and temporally). A landscape perspective to maintaining biodiversity has been prevalent in many early papers in ecology, natural history, and wildlife biology (Risser et al. 1984). The discipline of landscape ecology has gained recent prominence because it is an attempt to integrate a broad range of disciplines to study physical and ecological processes at the landscape-level. Ecology is defined as the study of the interactions among organisms and their environment (Dramstad et al. 1996). Forman (1998) defined a landscape as a kilometres-wide mosaic over which particular local ecosystems and land-uses recur and form a pattern. In a more biologically-oriented view, Maser (1993) defined a landscape as " a mosaic of habitat patches across which organisms move, settle, reproduce and eventually die". Thus, the discipline of landscape ecology examines both biotic and abiotic interactions that occur at the landscape scale. The management of total landscapes requires that we understand the theory and behaviour of ecosystems and how their processes are maintained and changed by natural and human-induced modifications (Barrett and Bohlen 1991). In recent decades a number of general principles have emerged in landscape ecology (Forman 1998). These principles include: (1) landscape structure and function, (2) biotic diversity, (3) species flow, (4) nutrient redistribution, (5) energy flow, (6) landscape change, and (7) landscape stability. The "patch-corridor-matrix" model (Forman 1998) has become a central component of landscape ecology in theory, and in practice. All landscapes are characterized by degrees of heterogeneity (patchiness) at different scales. Differing substrates (soils, bedrock), natural disturbances (fire, insect outbreaks), and human activity (forestry, road building) all create patchiness across a landscape. A patch is a homogenous area that differs from its surroundings (Forman 1998). Woodlots surrounded by farmland, or a wetlands immersed in upland habitat are examples of patches. Forman (1998) noted that patch shape correlates with the intensity of human activity. Intense human activity often results in simpler, less convoluted patch shape. Corridors are a form of patch in that they differ from the surrounding context; However they are usually identified as strips that aid in flows between patches (Lindenmayer 1994). Corridors fulfil a number of roles including: facilitating animal dispersal, wildlife habitat, preventing soil and wind erosion, and aiding in integrated pest management (Barrett and Bohlen 1991). A riparian buffer strip might serve as a corridor for forest song birds (Machtans et al. 1996), or kilometres-wide forested strip could serve as a corridor for cougar (Beier 1995). The life history traits of each species will determine the characteristics of corridor habitat. The matrix is the most extensive component of the landscape, is highly connected, and controls regional dynamics (Forman 1998). The Fundy Model Forest is characterized by a mixture of agricultural matrix (e.g. Kennebecasis Ecodistrict), and forest matrix (e.g. Anagance Ridge and Fundy Coastal Ecodistricts). Embedded the agricultural matrix are patches and corridors of forested land and human settlement. Within the forest matrix exist patches of recent clearcuts, human settlement, and a range of natural non-forest patches such as wetlands. The landscape structures briefly described above (patches, corridors, and matrix) influence, and are influenced by, landscape flows. These flows include a diversity of elements such as wildlife (Lindenmayer and Nix 1993), soil and nutrients (Stanley and Arp 1998), and water (Campbell 1970). For example, the flow of eastern newts (Notophthalmus viridescens) from natal wetland habitat to upland habitat is likely to be impeded by recent clearcuts (Waldick 1994). Thus, a matrix of farmland and clearcuts would reduce the flow of newts between habitat patches. One of the central principles of landscape ecology is that all ecosystems are interrelated, with movement or flow rate dropping sharply with distance, but more gradually between ecosystems of the same type (Forman 1998). Thus, a very heterogeneous landscape (with many patch types) is marked by a relatively low degree of movement (flow), and a large amount of resistance. All of these landscape components together create a landscape pattern or landscape structure. Landscape pattern thus refers to the number, size, and juxtaposition of landscape elements (Dunn et al. 1991). Landscape composition refers to the type of patches, corridors and matrix that exist in a landscape. Fragmentation is the breaking apart' of habitat. This can occur as a result of natural processes such as forest fires or anthropogenic disturbances such as road building or timber harvesting. Forman (1998) suggested that fragmentation is simply one phase of five in the land transformation process. Perforation occurs when holes are made in a habitat. For example, an extensive forest is perforated by logged areas or blowdowns. Dissection is the result of habitat being divided by equal-width lines such as roads, hydro-corridors or railway tracks. Fragmentation is the breaking up of habitat into smaller parcels. Shrinkage is the result of a decrease in the size of remaining patches or corridors. An example of this process is the declining area of woodlots in a farmland matrix that occurs as a result of housing development. Finally, attrition is the disappearance of patches and corridors. Jaeger (2000) added an additional important land transformation category called incision. This phase occurs when a linear element enters a habitat type, but does not separate the area into two patches (dissection). However, in the majority of the literature, fragmentation is most commonly referred to in a more general sense that is equivalent to Forman's (1998) entire five step process of land transformation. Different views exist on the relationship between habitat loss and fragmentation. Wilcove et al. (1986) stated that fragmentation is a combination of habitat loss and isolation. However, a number of ecologists have defined habitat loss and fragmentation separately, and have tried to analyze the independent effects of each on a range of organisms (Andren 1994, Fahrig 1998, Mazerole and Villard 1999). In these instances, fragmentation is defined purely as the breaking apart of habitat and does not imply habitat loss. Such approaches rely on various statistical techniques to separate the often confounding effects of habitat area and degree of fragmentation. In recent decades there has been an explosion of literature on the impacts of habitat fragmentation on forest species. The origin of much of this research can probably be traced to MacArthur and Wilson's (1967) "equilibrium theory of island biogeography". This theory, based on observations of island archipelagoes, related size and proximity to the mainland to species richness. Larger, closer islands tended to have greater numbers of species than smaller, more isolated islands. Isolated islands also tended to have higher rates of extinction than larger islands. In terrestrial ecosystems, fragmentation effects have been observed for birds (Verboom et al. 1991, Robinson et al. 1995, Estades and Temple 1999), mammals (Merriam 1988, Beier 1995, Hargis and Bissonette 1997), plants (Ouborg 1993, Schemske et al. 1994, Drayton and Primack 1996), amphibians (Petranka et al. 1994, Demaynadier and Hunter 1997), and insects (Roland and Taylor 1995, Haddad 1999). Alterations to landscape pattern and composition caused by fragmentation often have species-specific effects. An insurmountable barrier for one species, might be easily navigated by another. Landscape level fragmentation effects can be grouped into four major categories: patch size, distance between patches (connectivity), patch shape and edge. Edge Effect Edges are the result the result of vegetational boundaries in the landscape and may be caused by (1) enduring features (soils, drainage, slope), (2) natural disturbances, and (3) human activities such as clearcutting or farm development (Forman 1998). Edge effect is caused by differences in moisture, temperature and light that occur along the boundary between different adjacent patch types (Saunders et al. 1991). Edge zones may be characterized by high population density and diversity of species (Forman 1998). However, species that typically reside in edge habitat are usually relatively common. Species that require conditions associated with large tracts of mature, closed canopy forest are sometimes negatively affected by edges. For example, in tolerant hardwood woodland surrounded by agricultural matrix in Ontario, Burke and Nol (1998) found nine plant species that did not grow within 20 m of the forest edge. In an extensive study on avian reproductive success, Robinson et al. (1995) found that nest predation increased with decreasing forest cover. This was attributed primarily to the elevated amount of edge in fragmented landscapes and the preference of nest parasites (cowbirds) for edge habitat. Demaynadier and Hunter (1997) reported that salamander abundance is negatively affected by edge to distances between 25 and 35 m. Patch Size Many organisms are also affected by the size of favourable habitat patches. Such species are termed area sensitive (Freemark and Collins 1992). Robbins et al. (1989) found that "area" was one of the most significant habitat features for many neotropical migrant bird species. Robbins et al. (1989) generated a series of "predicted probability of occurrence" values for neotropical migrant species. For most birds, probability of occurrence increased with increasing forested area (Scarlet Tanager, Black-and-White Warbler (Mniotilta varia), Black-throated Blue Warber (Dendroica caerulescens)). Area sensitivity has also been observed for amphibians. Rosenburg and Raphael (1986 cited in Waldick 1994) found that amphibian species' densities were lower in patches smaller than 10 ha. While some debate exists about the area sensitivity of plants, a number of published studies report lower heterozygosity (genetic diversity) and higher rates of extinction in smaller populations (Bell et al. 1991, Damman and Cain 1998). Oubourg (1993) found that in a fragmented landscape, plants with poor dispersal capabilities are more likely to go extinct and less likely to recolonize habitat patches than highly vagile plants. Configuration: Connectivity and Distance Between Patches In some cases, fragmented landscapes have been shown to exhibit the same characteristics as those observed in island archpelagos by MacArthur and Wilson (1967). Isolation of habitat seems to compound the effect of small patch size on the ability of some species to persit and recolonize. These findings can be understood better if placed in the context of the concept of metapopulations'. The metapopulation concept requires that population dynamics be studied beyond the scale of local populations. Equilibrium', rather than occurring in a single, local population, might occur as a result of a number of inter-connected sub-populations that are distributed across a region (Husband and Barrett 1996). Population dynamics are the result of a series of local extinctions and recolonizations in habitat patches (Levins 1970). If the sub-population of one patch becomes extinct, it may eventually be recolonized by propagules from a sub-population that exists in a neighbouring patch. Husband and Barrett (1996) call this the "rescue effect". For a species to spread or persist, individuals must colonize unoccupied habitat patches as frequently as populations become extinct (Schemske et al. 1994). As fragmentation progresses, the distance between patches (isolation) of mature forest increases. This distance limits the ability of organisms to disperse and colonize new habitat patches. While very little empirical evidence exists, there is likely to be a threshold number of inhabited patches, and a critical distance between patches for each species, below which metapopulations begin to decline (Loo 1997). The term connectivity refers to the spatial continuity of a habitat patch type, matrix or network (Forman 1998). The matrix is inherently connected. However, a patch type will vary in connectivity depending on the proportion of the landscape that it occupies, the degree of patch clustering', and the existence of corridors. As noted above, corridors are strips that differ from their surroundings (the matrix) and provide opportunities for wildlife movement. Evidence is accumulating that corridors are used by a wide range of organisms for dispersal, migration, and daily movements within home ranges (Haas 1995, Husband and Barrett 1996, Beier and Noss 1998). By minimizing the distance between patches, corridors decrease patch isolation thus potentially reducing isolation of populations. Patch Shape Shape may have a critical effect on the ecological roles of patches. Forman (1998) outlined three "form-and-function" ecological principles of shape; (1) Compact forms are effective in conserving resources. Lower perimeter to area ratios reduce the amount of exchange between patch interior and the matrix, (2) Convoluted forms are effective in enhancing interactions with the surroundings. Longer boundaries in relation to patch interior provide a greater probability of movements across an edge, (3) Network forms tend to serve as a conduit system for transport. Forman (1998) provided the example of corridor-shaped patches such as a road or stream network facilitating movement (e.g. of people and sediment respectively). Patch shape may also provide an indication of degree of human impact on a landscape. In general, landscapes that have been greatly altered by resource exploitation or development tend to have patches with simpler shapes. Landscapes that are free of human influence tend to have more complex patch shapes (Krummel et al. 1987, Forman 1998). While studies showing the ecological relevance of patch shape are sparse, some evidence exists for the importance of this factor. Buechner (1989 cited in McGarigal and Marks 1995) reported that patch shape influenced small mammal migration. Hamazaki (1996) found that elongated and convoluted patches (in this case plywood boards) contained more common millipedes (Oxidus gracilis) than simply-shaped, more compact patches of the same area. The primary ecological effect of patch shape appears to relate to the edge effect. Patches that are long and narrow may be dominated entirely by edge. Circular patches will have comparatively more interior (Bender et al. 1998). It is important to note that in addition to the studies briefly described above that show significant influence landscape pattern on species distributions, there are many studies that reveal only weak or non-existent landscape effects (Game and Peterken 1984, Simberloff and Gotelli 1984, McGarigal and McComb 1995, Schmiegelow et al. 1997, Delin and Andren 1999). Using computer simulation, Fahrig (1997) found that the effects of habitat loss outweigh the effects of habitat fragmentation. Nevertheless, the precautionary principle' suggests that at the very least, in the face of uncertainty landscape changes that have the potential to affect native populations should be monitored. This is the domain of landscape metrics. 3.0 Landscape Metrics Landscape pattern and composition is highly variable. Landscape metrics are simply methods for quantifying this variability. With the use of landscape metrics it is possible to compare degree of fragmentation among landscapes (Tinker et al. 1998, Kitzberger and Veblex 1999) , or the rate of landscape change within a single landscape (Zheng et al. 1997). Jaeger (2000) listed five potential reasons for landscape quantification: (1) To document landscape development and validating observations, (2) To assess fragmentation within a region for comparison with other regions, (3) To research relationships between structural properties, landscape functions and the direction of landscape change, (4) To test hypotheses about the existence and location or type of critical thresholds in spatial pattern, and (5) To sharpen the conception of fragmentation by the discussion of quantitative expressions. Ecologists have grouped landscape metrics according to a number of different classification schemes. McGarigal and Marks (1995) made the important distinction between metrics of landscape composition (e.g. the proportion of area in different cover types), versus landscape configuration (e.g. shape, distribution, and area of patches). These authors and others (Baskent and Jordan 1995) have also emphasized the importance of scale in the landscape metrics. The measurement of landscapes is a hierarchical process involving several structural measurements at various scales. Measurements can be made for individual patches (e.g. size, shape, distance from neighbors), habitat class or cover type (e.g. patch size distribution, mean nearest neighbor), and landscapes (e.g. patch diversity, interspersion). Often class or cover type measures are simply the summary of patch-scale values. Baskent and Jordan (1995) also recommended that at each level, measurements be grouped into lineal, areal, and topological categories. Lineal measures involve the length, width and shape of patch boundaries. These relate directly to edge effect. Areal measures relate to the size of patches, or patch classes. The authors include patch shape in this category because it, along with edge effect, affects the core area of patches. Toploogical metrics are more relevant for the study of whole landscapes than individual patches. Such measures are concerned with the relative location of patches, corridors and the matrix (configuration). For the purposes of simplicity (and relevance), description of landscape metrics in this paper will relate each measure to the landscape fragmentation effects summarized above (patch size, patch shape, edge effect, and configuration). To reflect the importance of percent habitat cover in relation to independent fragmentation effects (Fahrig 1998), this will be put forward as a crucial fifth category of measurement.
Measuring the area of different habitat types is a simple but effective method for determining landscape change. As mentioned above, not all species require that suitable habitat be present in a single contiguous patch. Dunning et al. (1992) described a process called "landscape supplementation" in which the population in a focal patch may be increased if that patch is located in a portion of the landscape that contains additional available resources. Organisms might supplement their resource intake by using resources in nearby patches of the same habitat. For example, Barred Owls (Strix varia) which are normally restricted to larger patches have been found in a series of small patches in a fragmented woodland (Whitcomb et al. 1977). Many plants may not be sensitive to patch size or landscape configuration (Game and Peterken 1984). Landscape composition is typically described by (a) the number of categories or classes in a map and the area associated with each, (b) the proportion of each class relative to the entire map, and (c) diversity (Gustafson 1998). Diversity measures are influenced by two components: richness and evenness (McGarigal and Marks 1995). Richness refers to the number of patch types present; evenness refers to the distribution of area among different types. Many indices have been developed to measure landscape diversity, most of which are based upon measures previously used to determine species diversity (Shannon's diversity index and Simpson's diversity index). Area of habitat types is probably the most ecologically relevant of these three descriptors. Provided that time-series spatial data are available, with the use of area statistics it would be possible to determine how the area sensitive habitats such as Rich Northern Hardwood Forest or Hemlock Slope Forest (MacDougal and Loo 1996) have changed in the Fundy Model Forest over a certain time period. Diversity measures, while useful for monitoring landscape change within a region over time or among regions, are not relevant in isolation from other metrics. Variable patch type diversity may be the result of either human-induced or natural factors. For example, clearcut harvesting may convert a relatively homogenous landscape into a highly diverse, patchy landscape. Alternatively, a landscape typified by highly variable substrate and topography may exhibit a natural diversity that would be simplified by conversion to agricultural fields. Further, the use of diversity measures has been criticized because results convey nothing about the actual composition of a landscape (McGarigal and Marks 1995). A landscape may have high diversity, but be characterized by many non-indigenous patch types (e.g. urban area, non-native tree species plantations). 3.2 Patch Size Metrics If digital maps broken into patch categories are available, patch size is one of the simplest and most intuitive metrics. Mean patch size is often used to provide a summary of a particular patch type size (e.g. tolerant hardwood) within a landscape. However, averages may often be misleading due to the potential for skewed distributions. For example, the majority of patches in a landscape could be under 1 ha while a single contiguous patch might be 300 ha. Thus, the average patch size would not reflect the true landscape pattern. For this reason, many analysts recommend the use of patch size frequency distributions (Betts 1999), as well as patch size standard deviation, variance, and median (McGarigal and Marks 1995, Baskent and Jordan 1995). By examining the distribution of patch sizes it is possible to determine what percentage of patches fall within the spatial requirements of a particular species. For example, Woodley (1993) found that less than 20% of remnant forest patches in the Greater Fundy Ecosystem were larger than 50 ha. Because fisher (Martes pennanti), Pileated Woodpecker (Dryocopus pileatus), and Sharp-shinned Hawk (Accipiter striatus) all favour patches larger than this, these species are likely to be affected by this degree of fragmentation. If patch size metrics are used to quantify differences among landscapes, it is important that they be weighted to account for variance in the area of each landscape. McGarigal and Marks (1995) developed a series of area-weighted patch size metrics for this purpose. While patch size metrics may be ecologically relevant and relatively easy to implement, they may be problematic in landscapes where patches are not easily identifiable. Gustafson (1998) noted that many ecological and environmental conditions are characterized by gradients rather than discrete boundaries. GIS maps that delineate patches defined by single boundaries ignore these gradients. Further, definitions of what constitutes a patch may be purely arbitrary from a the perspective of a particular species. For example some salamander species might not discriminate between intolerant and tolerant hardwood forest (DeMaynadier and Hunter 1997). Nevertheless, in landscapes where recent human-induced change is prevalent, patches are often more easily identifiable. Recent clearcuts and agricultural fields are defined by distinguishable boundaries. In these cases, and cases where the habitat preferences of indicator species are well known and delineated, measurements of patch size are appropriate. 3.3 Edge Metrics Edge effect is highly variable and dependent upon habitat type, the composition of adjacent patches and the species under examination. Laurance and Yensen (1991) developed three steps to measuring edge effect for a patch type in a given landscape. First, appropriate local taxa must be identified. Such species should be among the most sensitive to edge effect. Second, the edge function' must be determined. This describes the response of a species to edge. Finally, Laurance and Yensen (1991) recommended the use of a core-area model to estimate the impact of edge on remaining habitat. Core area is defined as the total available habitat minus the edge effect (Fig. 1a). Examining core area is preferable to determining edge effect alone. Hargis et al. (1998) noted that edge density tends to increase with increasing representation of a disturbance cover type. However, as disturbance becomes more prevalent, edge density actually declines as remnant patches disappear (through attrition) and the landscape becomes more homogenous. As a consequence, both low and high-disturbance landscapes have similar edge values making it difficult to observe correlations between edge density and ecological phenomena over increasing disturbance (Hargis et al. 1998). Baskent and Jordan (1995) emphasized the importance of adjacent patch types in the determination of edge effect and core area. They stressed that the "vanishing distance" (the distance from a patch boundary inward to where edge effect is eliminated) can vary around the perimeter of a patch. Vanishing distance increases with elevated contrast between a patch and surrounding conditions (Fig. 1b). The shape of a patch also greatly influences the amount of core area. Narrow, elongated patches are less likely to have large core areas than patches that are compact in form. Similarly, patch shapes with irregular boundaries have greater edge effect than simple patch shapes (Hargis et al. 1998). Baskent and Jordan (1995) used a process called medial axis transformation (MAT) to account for these various influences of landscape context and patch shape on core area. Fig. 1 Edge effect and core area. The core area of the first patch (a) is reduced at a constant vanishing distance. The second patch (b) is reduced at variable distances due to different adjacent patch types. Edge 1 is adjacent to an immature forest stand. Edge 2 is adjacent to an agricultural field (after Baskent and Jordan [1995]). 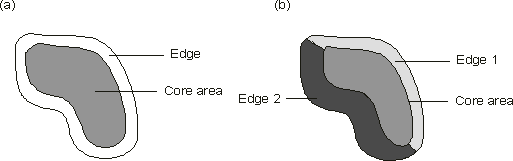
Nevertheless, if edge effect can be determined, with the use of GIS it is quite simple to determine the impact of this process on remaining habitat at the landscape level. It is often instructive to determine the proportion of total habitat area that is likely to be affected by edge influences. For example, based upon a range of edge effects reported in literature for herbaceous plants, Betts (1999) determined the amount of available core habitat remaining in the Saint John River Valley. 3.4 Landscape Configuration Metrics A wide variety of metrics has been used to measure the configuration of landscapes. Each of these attempt to reflect the degree to which patches are isolated or connected across landscapes. Nearest neighbour statistics are the most commonly cited configuration metric (McGarigal and Marks 1995, Baskent and Jordan 1995, Hargis et al. 1998). Mean nearest neighbour summarizes the distance between each patch of a certain cover type. This metric can be used along with information on the movement capabilities of various species to determine whether, on average, the distance between patches is close enough for organisms to interact (colonize, disperse, mate, migrate etc.). As in the case of mean patch size, mean nearest neighbour provides an incomplete picture of the spatial realities of a landscape. Properties such as clustering of patches can not be determined with the use of this metric alone (Rogers 1993 cited in Hargis et al. 1998). For this reason, McGarigal and Marks (1995) recommended the use of nearest neighbour standard deviation which shows the degree of variability about the mean. Proximity index has also been used to measure patch isolation (Hargis et al. 1998). Unlike nearest neighbor statistics, the proximity index is affected by patch size. This index is very sensitive to the spatial distribution of patches. Hargis et al. (1998) recommend the use of this statistic when patch isolation is comparatively high as in the case of gap analysis of species distribution and the study of spatial patterns of metapopulations. Baskent and Jordan (1995) proposed isolation as an effective measure of landscape configuration. Unlike nearest neighbour and proximity indices, isolation indicates how stands in a forest are different from conditions in their neighbourhood. Degree of isolation depends upon the scale of the neighbourhood. At a larger scale, isolation of patches across a landscape might increase or decrease. Contagion may also be used to measure landscape configuration (Li and Reynolds 1993). This metric has only been used for raster (pixel-based) maps. The contagion index is the sum over patch types of the product of two probabilities: (1) the probability that a randomly chosen cell belongs to a certain patch type i and (2) the conditional probability that given a cell is of patch type i, one of its neighbouring cells belongs to a different patch category (McGarigal and Marks 1995). Generally, a landscape with interspersed patch types will have lower contagion than a landscape with clustered patch types. McGarigal and Marks (1995) improved the contagion metric developed by Li and Reynolds (1993) to allow for analysis of patch (vector-based) adjacencies. They termed this the interspersion and juxtaposition index. The advantage of this approach is that it does not vary with image resolution (pixel size). Also, it may be applied to both to single cover types (classes) and a landscape as whole. Forman (1998) proposed a method for measuring the connectivity of landscapes. Corridor density is simply the number or abundance of corridors in an area. It is usually measured as total corridor length per unit area. Network connectivity was defined as the degree to which all nodes are connected (Forman 1998). This metric is the inverse of the proportion of linkages that must be added to have a connected system. However, very little evidence exists to answer the question of whether having all nodes connected is ecologically important. Thus such measures are mostly theoretical. Schumaker (1996) proposed patch cohesion as a potential measure of habitat connectivity of habitat as perceived by dispersing organisms. This measure, which can only be used on raster-based images, "connects" cells of the same cover type that share a common edge. In a similar vein, Jaeger (2000) developed measures of landscape configuration that are based on the ability of two animals placed in different areas somewhere in a region to find each other within the landscape. These metrics are rooted in the notion that interaction between populations is a precondition to the survival of many species. Jaeger (2000) proposed the use of three interacting metrics landscape division, splitting index, and effective mesh size to allow for comparisons of connectivity among landscapes. Landscape division is the probability that two randomly chosen places are not situated in the same undissected area. Splitting index is the number of patches one gets when dividing the total region into parts of equal size in such a way that this new configuration leads to the same degree of landscape division as in the original landscape. Effective mesh size reports the size of the patches created in the landscape generated by the splitting index. While less intuitive than other measures, these metrics are useful in that they allow for comparison among landscapes of different size. Lacunarity analysis of landscape patterns also provides information on the difficulty dispersers might have in locating suitable habitat (With and King 1999). This metric focuses on the gaps in habitat rather than the structure of habitat patches. Three major factors determine lacuniarity; (1) Gap size: landscapes in which the sizes of gaps are uniform at a particular scale have low lacuniarity values whereas landscapes with extremely variable gap sizes exhibit high lacuniarity values, (2) Proportion of habitat: landscapes with low proportions of habitat exhibit high lacuniarity values, (3) Spatial patterning of the landscape: higher lacuniarity values result from clumped habitat distributions than random or regular patterns (Plotnick et al. 1993). 3.5 Patch Shape Metrics Patch shape is probably the most difficult landscape feature to measure effectively. While a number metrics exist for this purpose, no proven accepted method exists. Forman (1998) suggested several methods for measuring patch shape. Patch elongation may be described by length and width ratio. Length and width are the dimensions of the narrowest rectangle that encloses a patch. To measure the convolution of a patch, Forman (1998) recommended counting the number of lobes. Lobes are protrusions that are longer than the radius of the largest circle that fits within a patch. The shape index represents the variation of a patch from a circle (Baskent and Jordan 1995). Fractal dimension measures the complexity of patches. Simple shapes such as circle and squares have fractal dimensions of 1. However, as shapes become more complex (boundaries become longer per unit area), fractal dimension approaches 2. FRAGSTATS uses this sort of analysis to measure patch shape at the landscape level. The landscape shape index summarizes the average patch characteristics for a particular cover type or the landscape as a whole. This index quantifies the amount of edge present in a landscape relative to what would be present in a landscape of the same size but with a simple geometric shape (McGarigal and Marks 1995). Measuring patch shape has proven to be problematic in that no single measurement or index of shape can unambiguously differentiate all shapes (Forman 1998). Further, depending on the resolution of an image, patch shapes may vary (McGarigal and Marks 1995). For example patches from a raster image with pixels representing 30 m2 on the ground exhibit more complex patch shapes than a vector-based GIS image. For these reasons, it is important to insure consistency in the resolution of maps to be compared. Further, it is most effective to choose the components of shape that are of ecological interest and then select metrics that can differentiate patches based on these components. 3.6 Summary A wide range of metrics exist for the measurement of habitat area, patch size, edge effect, configuration and patch shape. Only the most commonly used of these have been described above. As might be expected, many metrics substantially overlap. Interestingly, a number of these metrics, even in different categories, also co-vary (they are highly related). Ritters et al. (1999) examined 55 metrics using Principle Component Analysis. With the use of mapped landscapes from across the United States they determined that 87% of variation could be explained with the use of six metrics. These metrics were: average patch compaction' and image texture' (configuration metrics), patch-perimeter scaling' and average patch shape' (edge and shape metrics), number of attribute classes' (a habitat area or composition metric), and large patch density-area scaling' (a patch size metric). It is interesting to note that these authors did not examine metrics relating directly to connectivity or isolation. If species dispersal is a central focus of landscape analysis, it is likely that these metrics would contribute further explanatory power. An important message that is reiterated in much of the landscape metric literature is that no single metric is satisfactory for monitoring landscape change, examining the impacts of fragmentation on organisms, or comparing landscapes (Hargis et al. 1998, Ritters et al. 1999). Overall, metrics which are applicable at the class or cover-type level are probably the most ecologically relevant. Landscape metrics that attempt to summarize all landscape features simultaneously such as diversity indices and landscape interspersion are not useful except as a baseline for monitoring change within a single landscape. Further, the best metrics can be determined by the character of the landscape, the nature of the landscape change under examination, and the biological processes that are of interest. While in landscape ecology generalizations are often misleading, based on the literature, I have attempted to summarize the five metrics categories presented above by the way each would change for a mature forest-dependent species as a result of increasing human disturbance (Table 1). The following section summarizes several studies which have applied the range of metrics described above. Table 1. Generalized predicted direction of change as a result of human-induced disturbance for the five broad landscape metrics categories.
4.0 Application of Landscape Metrics Landscape metrics have been used in a variety of applications including determining the optimum spatial layout of timber harvest (Baskent 1999), assessing degree of landscape change from historical conditions (Hessburg et al. 1999), comparing natural versus human-altered landscapes (Krummel et al 1987, Kitzberger and Verblen 1999), and comparing degree of anthropogenic disturbance among watersheds (Wickham et al. 1999). The power of a landscape metrics approach depends upon the type of landscape(s) studied, the time scale across which landscape comparison is conducted, and the relevance of metrics to the ecology of identified species and processes. While studies that provide metric values without reference to other landscapes, past conditions, or the biological requirements of species may be of theoretical interest, they have little direct management relevance. For example, Sorrell (1998) used a suite of metrics to examine an agricultural/ forest landscape in the Thousand Islands region of southeastern Ontario. Sorrell (1998) measured a variety of patch, cover type and landscape level metrics including number of forest patches, total core area, mean core area, mean nearest neighbor, mean shape index, double log fractal, and several diversity indices. However all of these metrics were simply stated as absolute values. A mean shape index value of 1.44 means very little unless placed in the context of information about a species which indicates that it might require a minimum shape index value, or data from historical landscapes that allows for some sort of comparison. Sorrell (1998, 7) stated " it is believed that future research efforts should focus on species specific tolerances to fragmentation across a broader time and space horizon." One potential obstacle to performing landscape analyses on time series spatial data is the differing resolution of spatial data sets. Pixel size will affect several of the metrics described above. Shape metrics are most seriously effected. For this reason, Sorrell (1998) was not able to compare all metrics between historical and present landscapes. However, he did examine cover type area change between historical conditions (from 1977 air photos) and the present day. This valuable analysis made it possible to determine, for example, how deciduous forest has changed in relation to coniferous forest over a 20 year time span. Nikora et al. (1999) used area-length ratios, patch orientation, and fractal scaling to examine spatial heterogeneity at a range of scales caused by water movement. According to these researchers, different landscape properties, such as topography, vegetation, and soil exhibited different metric values. Krummel et al. (1987) used the perimeter-area method to determine the fractal dimension of deciduous forest patches in the Natchez Quadrangle a region characterized by recent conversion to cropland. Very different values were found for large patches in comparison small patches. The authors attributed this to the scale of human versus natural processes. Higher fractal dimension values were found if topographic and hydrological patterns were the major factors controlling forest patterns. Lower fractal dimension values were correlated with small patches of agricultural development. While both of these studies are valuable in that they show the responsiveness of particular metrics to natural and man-made landscape features, metric values themselves do not have much management relevance unless they can be related to thresholds associated with ecological processes at the landscape scale. Landscape metrics have been used to compare the degree of human-induced change among different landscapes. Tinker et al. (1998) analyzed landscape-scale pattern of cover types for 12 major watersheds in north-central Wyoming. By comparing landscape pattern metrics across remotely sensed GIS maps of watersheds, they determined the relative effects of clearcutting and road building on landscape pattern. Satellite images were classified into 24 cover types based on spectral reflectance values. Rather than analyzing individual metrics, Tinker et al. (1998) used Principal Component Analysis to group un-correlated metrics that explained most of the variation in landscapes. Metrics examined included area, edge, diversity, and shape. Interestingly, it was found that roads had a greater influence on fragmentation, as measured by metric clusters, than clearcuts. Tinker et al. (1998) suggested that the spatial arrangement of roads affected landscape structure than road density. To simplify calculations, these authors only examined cover type not seral stage. This is a serious drawback if measuring habitat values is a primary goal. A growing amount of research uses landscape metrics to examine changes in a single landscape over different time periods. Hulshoff (1995) analyzed landscape change in a Dutch farmland/ heath/ forest landscape using topographical maps from 1845 1982. A combination of what Hulshoff (1995) termed pattern indices' (patch number, patch size, and patch shape) and change indices' (change in % landscape type) were used. Changes in pattern indices were observed for time series as well. Hulshoff (1995) found that the average number of patches increased, while mean patch size declined. Two measures of patch shape were used, one of which was dependent on patch size (average perimeter area ratio), the other of which independent of this measure (core area in relation to core area of an isodiametric patch of the same area). Because trends in these metrics did not co-vary, Hulshoff (1995) recommended the use of several shape metrics to cover different aspects of ecological value. By examining change in cover type over the 167 year time span, it was possible for Hulshoff (1995) to determine periods that were characterized by the highest rates of change. One of the weaknesses of this study was that the metrics used provide no accurate estimation of the change in geographical position of land use over time. This is because no landscape configuration metrics were used. Hulshoff (1995) also stressed the need to attach ecological meaning to the index values that he generated. Kitzberger and Veblen (1999) quantified changes in fire frequency and associated vegetation pattern in mesic and xeric woodlands in Patagonia. Changes in landscape structure were examined with the use of digitized aerial photographs taken in 1940 and 1970. Rather than examining the entire landscapes, Kitzberger and Veblen (1999) relied on the use of sample areas. Declines in fire frequency have resulted in dramatic increases in forest cover in both coastal and inland areas of the study region. The majority of recently established forest has emerged from scrubland, while grasslands in each area have only changed minimally. Forest patches became more convoluted over the 30 year time span due to the expansion and fusion of formerly disjunct patches. Kitzberger and Veblen (1999) related many of these changes in landscape structure to the proximity of forest to shrubland patches. This is due to the dispersal abilities and regeneration success of native tree species. The strengths of this study lie in its time-series approach which allows the rate of change to be stated, and its examination of causal mechanisms that are related to the biology of local plant species. Zheng et al. (1997) used satellite imagery and landscape metrics to determine rates and patterns of landscape change as a result of forest harvest. Satellite images from 1972 and 1988 were examined for the Changbai Mountain area of China and Korea. FRAGSTATS (McGarigal and Marks 1995) was used to examine landscape structure and composition. However, Zheng et al. (1997) focused on three main landscape indices: edge density (the sum of edge divided by the total landscape area), patch density, and mean patch size. Amount of interior forest', defined as amount of forest remaining after designating a 100m edge zone. By all measures, forest harvesting accelerated over the study time span. Percent forest cover declined, number of patches increased, and edge increased. The strength of this study was that it established unambiguous trends in the extent of forest removal. However, because no individual cover types were identified, it could not be determined which habitats are under the most threat. Further, beyond estimating that " these changes will have a significant impact on composition of flora and fauna", trends were not related to ecological processes or the habitat requirements of local species. The authors concluded with recommendations for habitat protection based on coarse estimates (based on elevation) about relative threat to different habitat types. One of the most interesting studies that utilizes landscape metrics attempted to determine change in a forested landscape from a hypothetical reference' landscape. Hessburg et al. (1999) examined landscape pattern and composition for two time periods. "Historical" (1938-1956) and "current" (1985-1993) conditions were determined with the use of aerial photos. In addition, these researchers estimated the variation in pattern and structure that would have occurred in a reference condition. Reference variation, derived from the "historical" landscape, was used to determine the importance of spatial pattern change. Reference variation was set at 80% of the average values for the 343 sub-watersheds. The justification for this value was that the majority of landscape metric values tended to cluster around 80% variation. Ten landscape metrics were used to display spatial pattern conditions. These included diversity indices, configuration metrics (contagion and interspersion) and edge metrics (area-weighted mean edge contrast index). Hessburg et al. (1999) then tested this approach by conducting a spatial analysis on a sample watershed to determine how it varied from reference conditions. This approach is of value to land managers as a tool to determine if pattern changes that result from current management fall outside of the variation that would have existed within a historical landscape. This potentially defines a range (albeit an imperfect range) of ecologically justifiable management decisions. To summarize, many studies have used landscape metrics to quantify spatial pattern both among landscapes and within single landscapes. The most rigorous studies are those that determine landscape change based on time-series spatial data (historical versus current conditions). In order to avoid so what?'-type results, and to allow for natural variation and landscape change, it is important to establish a reference condition and an acceptable range of landscape pattern variation for management purposes (Hessburg et al. 1999). Of utmost importance is the relation of metrics to ecological processes and the life histories of native species. While a number of research projects have used metrics for comparative purposes, very few have successfully related the quantitative results provided by metrics directly to species requirements. 5.0 The Relative Importance of Landscape Pattern to Selected Taxa This literature review has emphasized the importance of relating landscape metrics to the ecological requirements of native species. One fundamental generalization in landscape ecology appears to be that when it comes to the impacts of fragmentation on organisms, no simple interspecific rules exist. What might constitute a fragmented landscape for one species (e.g. yellow spotted salamander [Ambystoma maculatum]), might be a highly connected landscape for another (e.g. White-throated Sparrow [Zonotrichia albicollis]). This section provides a brief analysis of the relation of the metrics described in Section 3 to the results of scientific studies on the spatial requirements of a range of forest-dependent taxa. It is hoped that this analysis will assist the Fundy Model Forest in selecting ecologically relevant metrics for landscape quantification. As discussed in Section 2, over the past decade a large body of literature has emerged on the effect of landscape pattern and composition on forest species. Studies from a wide range of ecosystem types have examined the effect of patch size (Weinburg and Roth 1998, Roland 1993), edge effect (Matlack 1994, Laurance 1991), total forest cover (AndrJn 1994), patch shape (Hamazaki 1996), connectivity (Beier and Noss 1998, Haas 1995), isolation (Opdam et al. 1994), and a range of other landscape configuration variables. Several reviews have been conducted on various landscape pattern and composition effects. Bender et al. (1998) quantitatively reviewed the results of 25 published studies that tested the relationship between patch size and population density. They found that patch size effects were strong for both edge species (positive effect for small patches) and interior species (positive effect for large patches), however generalist species exhibited negligible effects. Interestingly, no evidence was found which indicated that these patch size effects were related to landscape characteristics such as proportion of landscape covered by habitat, median patch size or the scale at which studies were conducted. However, it is likely that edge effect contributes greatly to patch size effect. Bender et al. (1998) were not able to separate these interacting variables. Nevertheless, they concluded that habitat loss and fragmentation affecting patch size will greatly influence the abundance of edge and interior species. The random sample hypothesis states that if habitat area is held constant, there should not be lower densities of species in suitable habitat of fragmented landscapes than in suitable habitat of contiguous landscapes. That is, species abundance is only affected by habitat loss, not fragmentation effects. AndrJn (1994) reviewed studies which tested the random sample hypothesis for birds and mammals. AndrJn (1994) found that there might be a threshold in proportion of suitable habitat in the landscape above which habitat loss is the only impact on species decline, but below which species decline is greater than would be expected as a result of habitat loss alone. He suggested that this threshold might be between 10% and 30% of suitable habitat. When fragmentation reaches these levels AndrJn (1994) showed accelerated declines in patch size and increases in patch isolation. Thus, while AndrJn's (1994) hypothesis is stated in terms of proportion of suitable habitat, the effects of this factor are likely to be governed by two other measurable landscape criteria: patch size and patch isolation. Beier and Noss (1998) reviewed 32 studies that tested the functionality of wildlife corridors. The authors dismissed many of the poorly designed studies. Nevertheless, they argued that the balance of evidence indicates that corridors are a valuable conservation tool. Thus, metrics that are capable of measuring this landscape feature should be seriously considered by the FMF. The only drawback to measurement of connectivity is the necessity to determine this feature on a species-by-species basis. As has been emphasized, due to varying habitat preferences and life histories, organisms may perceive a landscape in many different ways and at different scales. It is critical to ask the question: "connected for which species?" before applying connectivity metrics. Opdam (1991) reviewed avian research on metapopulation dynamics and habitat fragmentation. He found that a few studies based on time series demonstrate that local extinction is related to fragment size, while recolonization rate depends on the degree of patch isolation. Further, density of corridors' lowering landscape resistance also increased the probability of occurrence for some species. However, most of the studies examined were conducted in agricultural landscapes. Opdam (1991) highlighted the need for studies in less predictable habitat types. Mazerolle and Villard (1999) reviewed 61 studies that examined the effects of both landscape context and patch characteristics as predictors of species presence. They defined landscape context as both configuration' (e.g. isolation, nearest patch, connectedness) and cover' (e.g. amount of suitable habitat within a given distance). Patch characteristics included area, age, habitat, orientation, perimeter, perimeter/ area, and shape. Mazerolle and Villard (1999) found that patch variables were significant predictors of species abundance in 93.4% of the studies examined. Landscape and patch variables effectively predicted abundance in 52.5% of studies. Vertebrates were the most sensitive to landscape context (79.5% of studies reported a significant effect). Of the patch variables, patch size and habitat type were by far the most frequent explanatory variables. One study (McCollin 1993) reported that patch shape was an important factor. Further, only one study (Bellamy et al. 1996) reported that patch perimeter was a significant variable, however perimeter/ area ratio was a significant factor in four of the 61 studies examined. Of the two landscape variables considered, configuration' was an explanatory variable in more of the studies (37.7%) than cover' (31.1%). AndrJn (1994) reviewed the effects of landscape composition on predation rates at habitat edges. Overall, studies that reported significant edge effects in forest mosaics were far less frequent than those reporting edge effects in forest/ farmland landscapes. Paton (1994) reviewed the literature on the effects of edge on avian nest success. Despite methodological differences in studies on edge effect, the most conclusive studies showed effects within 50 m of a hard edge. Only one study was found that explicitly set out to test the relation of landscape metrics to the structure of biotic communities. Miller et al. (1997) computed a range of measures of land cover including: diversity, dominance, contagion, fractal dimension, mean patch size, forest edge, clustering of selected forest types, and size of largest forest patches, within two 100 km2 watersheds in Pennsylvania. The analysis was conducted at the sub-watershed level and sought to determine the influence of human disturbance on bird and vascular plant guilds. In general, the landscape metrics which corresponded to differences in human land use were the ones which related to differences in number of species. Of the metrics listed above, the general descriptors of diversity, contagion, mean patch size, proportion of forest cover, and amount of forest edge were the most effective at reflecting disturbance and changes in guild composition.
Overall, this summary of previous reviews of the influence of landscape structure on ecological processes and biotic communities indicate that a wide range of metrics may explain the distribution and abundance of species. Total proportion of suitable habitat and patch size seem to be the most frequently cited important landscape features. However, configuration metrics (e.g. connectivity, isolation, and contagion) are frequently reported as being significant features. A comprehensive survey of all literature on landscape pattern and composition is beyond the scope of this paper. A brief review of important research was provided in Section 2. However, in order to assess the relative importance of different landscape factors literature on the effects of landscape pattern and composition on species' habitat was gathered. Due to the time constraints of this study, not all of the literature on landscape effects was examined. Recent literature (within the past five years) was more intensively searched. Search emphasis was also placed on journals that frequently publish articles on landscape effects: Biological Conservation, Conservation Biology, Ecology, Ecological Applications, Ecological Monographs, Journal of Wildlife Management, Landscape Ecology, Oikos, Oecologia, and The Auk. Only studies showing significant landscape effects are reported. While a range of taxa were considered in this analysis (birds, amphibians, mammals, plants and insects), the majority of 36 studies reported focus on birds. Results should be put in the context this taxonomic bias. Further, it should be noted that the existence of a large number of studies that confirm a particular landscape effect may not indicate that this is the most significant factor. Rather this might reflect the ease with which this effect can be studied. This summary makes no attempt to weight studies based on the degree of statisical or biological significance in results. Table 2. Summary of studies reporting landscape effects. For each study, I report the major landscape effect(s) found (% cover, patch size, patch shape, edge, and configuration), metric types used (if any), and the biological variable affected. All results presented are statistically significant for at least one of the species under examination.
|
||||||||||||||||||
| Birds | Agricultural/ forest | Patch size, configuration | Patch size, nearest neighbor | Abundance | Villard et al. (1995) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Agricultural/ forest | Patch size, configuration | Patch size, isolation | Density | Matthysen (1999) | ||||||||||||||
| Agricultural/ forest | Patch size, configuration | Patch size, nearest neighbor | Extinction/ re-settlement rates | Verboom et al. (1991) | ||||||||||||||
| Grassland | Patch size | Patch size | Nesting success | Winter and Faaborg (1999) | ||||||||||||||
| Forest | Patch size | Patch size | Pairing success | Hagan et al. (1996) | ||||||||||||||
| Forest | Patch size | Stand width (riparian) | Presence/ absence | Kilgo et al. (1998) | ||||||||||||||
| Agricultural/ forest | Patch size | Patch size | Reproductive success | Weinberg and Roth (1998) | ||||||||||||||
| Agricultural/ forest | Patch size | Patch size | Presence/ absence | Robbins et al. (1989) | ||||||||||||||
| Agricultural/ forest | Patch size | Patch size | Reproductive success | Donovan et al. (1995) | ||||||||||||||
| Agricultural/ forest | Patch size | Patch size | Pairing success | Porneluzi et al. (1993) | ||||||||||||||
| Agricultural/ forest | Edge | NA | Nest predation | Donovan et al. (1997) | ||||||||||||||
| Forest | Configuration | Isolation, connectivity | Abundance/ diversity | Schmiegelow et al. (1997) | ||||||||||||||
| Forest | Configuration | Distance to roads | Pairing success | Ortega and Capen (1999) | ||||||||||||||
| Forest | Configuration | Connectivity | Movement | Machtans et al. (1996) | ||||||||||||||
| Agricultural/ forest | Configuration | Connectivity | Movement frequency | Haas (1995) | ||||||||||||||
| Forest | % Habitat cover | % Habitat cover | Presence/ absence | Drolet et al. (1999) | ||||||||||||||
| Forest | % Habitat cover, configuration | PCA^ | Presence/ absence | Trzcinski et al. (1999) | ||||||||||||||
| Forest | % Habitat cover, configuration | PCA^( | Presence/ absence | McGarigal and McComb (1995) | ||||||||||||||
| Agricultural/ forest and forest | % Habitat cover, configuration | PCA^(( | Presence/ absence | Rosenburg et al. (1999) | ||||||||||||||
| Agricultural/ forest | % Habitat cover + configuration | NA | Reproductive success | Porneluzi and Faaborg (1998) | ||||||||||||||
| Forest | % Habitat cover | % Cover | Reproductive success | Ripple et al. (1991) | ||||||||||||||
| Agricultural/ forest | % Habitat cover | % Cover | Reproductive success | Robinson et al. (1995) | ||||||||||||||
| Taxon | Landscape type | Landscape effect | Metric type | Biological variable effected | Reference | |||||||||||||
| Amphibians | Lake islands | Patch size | Patch size | Presence/ absence | Hager (1998) | |||||||||||||
| Forest | Edge | Edge distance | Density | DeMaynadier and Hunter (1997) | ||||||||||||||
| Forest/ residential | Configuration and edge | Road density, edge | Presence/ absence | Gibbs (1998) | ||||||||||||||
| Forest streams | Configuration | Isolation | Antipredator behavior | Storfer and Sih (1998) | ||||||||||||||
| Forest/ pond | Configuration | Isolation | Presence/ absence | Gulve (1994) | ||||||||||||||
| Agricultural/ urban/ forest | % Habitat cover | % Cover | Presence/ absence | Gibbs (1998) | ||||||||||||||
| Taxon | Landscape type | Landscape effect | Metric type | Biological variable effected | Reference | |||||||||||||
| Plants | Agricultural/ forest | Edge | Edge distance | Alien plant invasion | Brothers and Spingarn (1991) | |||||||||||||
| Agricultural/ forest | Edge | Edge distance | Presence/ absence | Burke and Nol (1998) | ||||||||||||||
| Agricultural/ forest | Edge | Edge distance | Presence/ absence | Matlack (1994) | ||||||||||||||
| Forest | Edge | Edge distance | Reproductive success | Jules (1998) | ||||||||||||||
| Agricultural/ forest | Patch size/ configuration | Connectivity | Species richness | Grashof-Bokdam (1997) | ||||||||||||||
| Grassland | Configuration | Isolation | Extinction/ colonization | Ouborg (1993) | ||||||||||||||
| Agricultural/ forest | Configuration | Isolation | Genetic diversity | Damman and Cain (1998) | ||||||||||||||
| Taxon | Landscape type | Landscape effect | Metric type | Biological variable effected | Reference | |||||||||||||
| Mammals | Forest | Edge | Edge distance | Presence/ absence | Mills (1994) | |||||||||||||
| Agricultural/ forest | Patch size | Patch size | Presence/ absence | van Apeldoorn et al. (1994) | ||||||||||||||
| Agricultural/ forest | Configuration | Connectivity | Density | La Polla and Barrett (1993) | ||||||||||||||
| Forest | % Habitat cover | %Cover | Presence/ absence | Hargis and Bissonette (1997) | ||||||||||||||
| Forest | % Habitat cover, patch size, configuration | % Habitat cover, Mean patch size, patch density | Presence/ absence | Reunanen et al. (2000) | ||||||||||||||
| Taxon | Landscape type | Landscape effect | Metric type | Biological variable effected | Reference | |||||||||||||
| Insects | Plywood board | Patch shape | Perimeter-area ratio | Density | Hamazaki (1996) | |||||||||||||
| Forest | Edge | Edge length | Duration of tent caterpillar outbreak | Roland (1993) | ||||||||||||||
| Agricultural/ forest | Configuration | Connectivity | Movement | Hill (1995) | ||||||||||||||
| Forest | % Habitat cover | % Habitat cover | Presence/ absence | Roland and Taylor (1997) |
Based on the studies listed above (Table 2), it is evident that a wide range of landscape effects have been reported for birds, amphibians, plants, mammals and insects. Overall, configuration (55.5%), patch size (39%), and percent habitat cover (30.5%) are the most frequently cited effects. However certain effects seem to be more common within each of the taxa examined. For example, most of the landscape-level bird studies reported patch size as an effective predictor of presence/absence, density or reproductive output. On the other hand, amphibians seem to be most commonly affected by configuration, although the most convincing study (Gibbs 1998) reported percent cover as the most important factor in determining the presence of certain amphibian species. Edge effect was reported as significant in four of the six plant studies which found landscape effects. The only landscape effect described in this report that was not commonly cited in the literature was patch shape.
The conclusion that can be drawn from both previous literature reviews, and the review conducted in this report is that a range landscape features (effects) are important and should be the focus for analysis in the Fundy Model Forest. Effects of particular importance are: (1) percent habitat cover, (2) patch size, (3) edge effect, and (4) configuration. This conclusion must be understood within the context of several caveats. First, effects vary in importance depending on the species of interest. Key, or indicator species should be selected before landscape analysis is conducted. Second, metrics should be selected on the basis of how directly they measure the landscape feature of interest. Less intuitive measures such as splitting index and effective mesh size (Jaeger 2000) may show correlation with species presence/absence or reproductive success, but are less likely to provide insight into causal mechanisms.
6.0 Conclusion and Recommendations: Application of Fragmentation Metrics in the Fundy Model Forest
This report has summarized the current literature on landscape metrics as they relate to the concepts of landscape ecology and biodiversity. From this survey of proposed landscape metrics, their application, and their relation to the requirements of native species, it is possible to propose a number of recommendations for the measurement of fragmentation in the Fundy Model Forest.
-
6.1 Metric Prioritization with an Indicator Species Approach
- Percent habitat cover
- Patch size distribution
- Edge effect
- Landscape configuration
- Principal metrics. These metrics should be as simple as possible and directly related to the habitat requirements of indicator species. Habitat area and patch size metrics are likely to fall into this category.
- Secondary metrics. These metrics will be used to triangulate' the results of the metrics used above. For example, it may be necessary to determine connectivity through several methods. While these metrics may not relate as directly to indicator species requirements, they will provide a means to check results of the principle metrics.
- Baseline monitoring metrics. These metrics may not have direct ecological relevance to indicator species, however they will provide baseline data on landscape structure change. As new knowledge becomes available on the spatial requirements of species, data on these metrics may become valuable. Metrics such as diversity, interspersion and patch shape are likely to fall into this category.
- Establish indicator species.
- Develop historical' condition and acceptable reference variation.
- Implement metrics for historical and present landscapes to determine rates of landscape change.
- Test the influence of habitat composition and pattern by monitoring actual populations of indicator species.
No single metric is capable of reflecting the diversity of landscape composition and pattern. However, four general categories of metrics should be central to any analysis of fragmentation in the FMF:
As was evident in the previous section, it is not possible to prioritize this list for all species because the importance of each metric set will vary depending upon the life history of the organism of concern. For this reason, it is recommended that an indicator species approach be adopted. Indicator species represent a larger group of organisms that exist in similar habitats. By managing for the needs of indicator species, the assumption is made that other species with similar habitat requirements will be protected (Nilsson et al. 1995). Generally, as it is older forest that is under threat from development, indicator species are characteristically old forest dwellers. The best indicators are also sensitive to a range of landscape effects such as patch size and configuration. To meet these criteria, it will be necessary to identify several indicator species that represent a variety of habitat types and have a range of dispersal mechanisms and patch size sensitivities. It is important to note that providing habitat for indicator species does not necessarily ensure protection of overall diversity (Flather et al. 1997). Detailed monitoring of a range of species must be undertaken to support the assumptions associated with an indicator species approach.
Once indicator species have been selected it is recommended that potential metrics be grouped into three major prioritized categories:
6.2 Methods and Tools
As has been discussed above, studies that do not put metrics in the context of (a) the habitat requirements of species, (b) changes from historical landscapes, or (c) comparison with other landscapes, have little management relevance. The most rigorous and useful studies establish some sort of baseline condition and acceptable variation in landscape pattern and composition (e.g. Hessburg et al. 1999). Adherence to the following four criteria will assist in developing a landscape fragmentation measurement approach that is relevant to forest management and biodiversity conservation in the Fundy Model Forest:
To add further power to this landscape analysis, it would be instructive to predict future landscape change. With the use of spatial forest management plans for industrial forest and stochastic modelling for private woodlots, it would be possible to estimate key metrics twenty years from now. A joint project of the Fundy Model Forest and Greater Fundy Ecosystem Group will attempt to develop such an approach*.
To conduct an landscape fragmentation analysis several tools are necessary: (1) An up-to-date Geographic Information System with age class and cover type delineation, (2) A spatial analysis and statistics program. In the past, such programs have been quite technically difficult, however recent developments have provided user-friendly interfaces. Several of the metrics summarized in this report are not included in such packages, however it is possible to either find or develop simple computer programs (ArcView scripts [ESRI 1998]) that could conduct these analyses, (3) If future projections or establishment of reference conditions are desired, a tool for modelling landscape change will be necessary. A number of computer-based tools, both stochastic and deterministic, have been developed (Fall 1999, Mladenoff and He 1999, Gustafson and Crow 1999).
To conclude, it is recommended that the Fundy Model Forest, in co-operation with all major landowners, initiate a landscape-level monitoring program. A wide range of potential metrics have been suggested in this report. Of utmost importance is that these metrics be used thoughtfully with regard for the habitat requirements for local indicator species, and placed in the context of historical landscapes and future landscape change. Results will then be relevant to forest management planning.
Literature Cited
AndrJn, H. 1994. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71: 355-366.
AndrJn, H. 1995. Effects of landscape composition on predation rates at habitat edges. In Mosaic Landscapes and Ecological Processes, Hansson, L. L. Fahrig, and G. Merriam (Eds.) London: Chapman and Hall.
Barrett, G. and P. Bohlen. 1991. Landscape ecology. In Landscape Linkages and Biodiversity. Hudson, W. (Ed.) Washington D.C.: Island Press.
Baskent, E. 1999. Controlling spatial structure of forested landscapes: a case study towards landscape management. Landscape Ecology 14: 83-97.
Baskent, E. and G. Jordan. 1995. Characterizing spatial structure of forest landscapes. Canadian Journal of Forest Research 25: 1830-1849.
Beier, P. and R. Noss. 1998. Do habitat corridors provide connectivity? Conservation Biology 12(6): 1241-1252.
Beier, P. 1995. Dispersal of juvenile cougars in fragmented habitat. Journal of Wildlife Management. 59: 41-67.
Bell, G., J. Martin, and D. Schoen. 1991. The ecology and genetics of fitness in forest plants III. Environmental variance in natural populations of Impatiens pallida. Journal of Ecology 79: 697-713.
Bender, D., T. Contreras, and L. Fahrig. 1998. Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79(2), 517-533.
Betts, M. 1999. A landscape ecological approach to conserving the Appalachian Hardwood Forest. Fredericton: Nature Trust of New Brunswick. Pp. 95.
Burke, M., and E. Nol. 1998. Edge and fragment size effects on the vegetation of deciduous forests in Ontario, Canada. Natural Areas Journal 18: 45-53.
Buford, E. and D. Capen. 1999. Abundance and productivity of forest songbirds in a managed, unfragmented landscape in Vermont. Journal of Wildlife Management 63(1): 180-188.
Brothers, T. and A. Spingarn. 1992. Forest fragmentation and alien plant invasion of central Indiana old-growth forests. Conservation Biology 6(1): 91-100.
Campbell, C. 1970. Ecological implications of riparian vegetation management. Journal of Soil and Water Conservation 2: 49-52.
Darveau, M., L. Beauchesne, L. Belanger, J. Huot, and P. LaRue. 1995. Riparian forest strips as habitat for breeding birds in boreal forest. Journal of Wildlife Management 59: 67-78.
Damman, H. and M. Cain. 1998. Population growth and viability analyses of the clonal woodland herb, Asarum canadense. Journal of Ecology 86: 13-26.
Demaynadier, P., and M. Hunter. 1997. Effects of silvicultural edges on the distribution and abundance of amphibians in Maine. Conservation Biology 12(2): 340-352.
Denlin, A. and H. AndrJn. 1999. Effects of habitat fragmentation on Eurasian red squirrel (Sciurus vulgaris) in a forest landscape. Landscape Ecology 14(1): 67-72.
Donovan, T., P. Jones, E. Annand and F. Thompson. 1997. Variation in local-scale edge effects: mechanisms and landscape context. Ecology 78(7): 2064-2075.
Dramstad, W., Olson, J., and R.T.T. Forman. 1996. Landscape Ecology Principles in Landscape Architecture and Land-Use Planning. Washington: Island Press
Drolet, B., A. Desrochers, and M-J. Fortin. 1999. Effects of landscape structure on nesting songbird distribution in a harvested boreal forest. The Condor 101: 699-704.
Dunning, J., B. Danielson, H. Pulliam. 1992. Ecological processes that affect populations in complex landscapes. Oikos 65(1): 169-175.
ESRI. 1998. ArcView 3.1.
Etheridge, P., K. Pugh, G. Watt, M. Whelan. 1999. Criteria and Indicator Status Report 1999/2000. Natural Resources Canada.
Estades, C. and S. Temple. 1999. Deciduous-forest bird communities in a fragmented landscape dominated by exotic pine plantations. Ecological Applications 9(2): 573-585.
Fahrig, L. 1998. When does fragmentation of breeding habitat affect population survival? Ecological Modelling 105: 273-292.
Fahrig, L. 1997. Relative effects of habitat loss and fragmentation on population extinction. Journal of Wildlife Management 61(3): 603-610.
Fall, A. 1999. SELES: A tool for modeling landscape structure and change. Unpublished information report. Vancouver: Simon Fraser University.
Flather, C., K. Wilson, D. Dean, and W. McComb. 1997. Identifying gaps in conservation networks: Of indicators and uncertainty in geographic-based analyses. Ecological Applications 7(2): 531-542.
Forman, R.T.T. 1998. Land Mosaics: The Ecology of Landscapes and Regions. Cambridge: Cambridge University Press.
Freemark, K. and B. Collins. 1992. Landscape ecology of birds breeding in temperate forest fragments. In Ecology and conservation of Neotropical migrant landbirds (Hagan, J. and D. Johnson (Eds.) Washington: Smithsonian Institution Press, Pp. 443-454.
Game, M., and Peterken, G. 1984. Nature reserve selection strategies in the woodlands of central Lincolnshire, England. Biological Conservation 29: 157-181.
Gibbs, J. 1998. Distribution of woodland amphibians along a forest fragmentation gradient. Landscape Ecology 13: 263-268.
Gill, D. 1978. The metapopulation ecology of the red-spotted newt. Ecological Monographs 48: 145-166.
Godt, M., B. Johnson and J. Hamrick. 1996. Genetic diversity and population size in four rare southern Appalachian plant species. Conservation Biology 10(3): 796-805.
Grashof-Bokdam, C. 1997. Forest species in an agricultural landscape in the Netherlands: Effects of fragmentation. Journal of Vegetation Science 8: 21-28.
Gustafson, E. and T. Crow. 1999. HARVEST: linking timber harvesting strategies to landscape patterns. In Spatial Modeling of Forest Landscape Change: Approaches and Applications. Mladenoff, D. and W. Baker (Eds.), Cambridge: Cambridge University Press.
Gustafson, E. 1998. Quantifying landscape spatial pattern: what is the state of the art? Ecosystems 1: 143-156.
Gulve, P.S. 1994. Northern metapopulation of the pool frog. Ecology 75(5): 1357-1367.
Haas, C. 1995. Dispersal and use of corridors by birds in wooded patches on an agricultural landscape. Conservation Biology 9(4), 845-854.
Haddad, N. 1999. Corridor and distance effects of interpatch movements: a landscape experiment with butterflies. Ecological Applications 9(2): 612-622.
Hamazaki, T. 1996. Effects of patch shape on the number of organisms. Landscape Ecology 11(5): 299-306.
Hargis, C., J. Bissonette, and J. David. 1998. The behaviour of landscape metrics commonly used in the study of habitat fragmentation. Landscape Ecology 13: 167-186.
Hargis, C. and J. Bissonette. 1997. Effects of forest fragmentation on populations of American marten in the intermountain west. In Martes: Taxonomy, Ecology, Techniques, and Management. Proceedings of the Second International Martes Symposium. Proulx, g., H. Bryant, and P. Woodard (Eds.) Edmonton: Provincial Museum of Alberta, Pp. 437- 451.
Hessburg, P., B. Smith, and R. Salter. 1999. Detecting change in forest spatial patterns from reference conditions. Ecological Applications 9(4): 1232-1252.
Hill, C.J. 1995. Rain forest corridors and insect dispersal. Conservation Biology. 9(6)1559-1565.
Hulshoff, R. 1995. Landscape indices describing a Dutch landscape. Landscape Ecology 10(2): 101-111.
Husband, B. and S. Barrett. 1996. A metapopulation perspective in plant population biology. Journal of Ecology. 84: 461-469.
Jaeger, J. A. 2000. Landscape division, splitting index, and effective mesh size: new measures of landscape fragmentation. Landscape Ecology 15: 115-130.
Jules, E. 1998. Habitat fragmentation and demographic change for a common plant: Trillium in old-growth forest. Ecology 79(5): 1645-1656.
Kilgo, J., R. Sargent, B. Chapman, and K. Miller. 1998. Effect of stand width andadjacent habitat on breeding bird communities in bottomland hardwoods.Journal of Wildlife Management 62(1): 72-83.
Kitzberger, T. and T. Veblex. 1999. Fire-induced changes in northern Patagonian landscapes. Landscape Ecology 14: 17-33.
Krummel, J. , R. Gardner, G. Sugihara, R. O'Neill, and P. Coleman. 1987. Landscape patterns in a disturbed environment. Oikos 48: 321-324.
La Polla, V. and G. Barrett. 1993. Effects of corridor width and presence on the population dynamics of the meadow vole (Microtus pennsylvanicus). Landscape Ecology 8(1): 25-37.
Laurance, W. and E. Yensen. 1991. Predicting the impacts of edge effects in fragmented habitats. Biological Conservation 55: 77-92.
Levins, R. 1970. Some Mathematical Questions in Biology 2: 75-107. Providence R.I. American Mathematical Society.
Li, H. and J.F. Reynolds. 1993. A new contagion index to quantify spatial patterns of landscapes. Landscape Ecology 7:101-110.
Lindenmayer, D. 1994. Wildlife corridors and the mitigation of logging impacts on fauna in wood-production forests in south-eastern Australia: a review. Wildlife Research 21: 323-340.
Lindenmayer, D., and Nix, H. 1993. Ecological principles for the design of wildlife corridors. Conservation Biology 7(3): 627-630.
Loo, J. 1997. Biodiversity as a forest landscape value: how can we maintain it? Canadian Council on Ecological Areas, Fredericton Workshop Proceedings, 87-91.
MacDougall, A. and J. Loo. 1996. Fine-Scale Community Types of the Fundy Model Forest in Southeastern New Brunswick. Canadian Forest Service Atlantic Forestry Centre, Natural Resources Canada, Information Report M-X-198E.
Machtans, C., M. Villard, and S. Hannon. 1996. Use of riparian buffer strips as movement corridors by forest birds. Conservation Biology 10(5): 1366-1379.
Maser, C. 1993. Adaptable landscapes are the key to sustainable forests.
Matlack, G. 1994. Vegetation dynamics of the forest edge trends in space and successional time. Journal of Ecology 82: 113-123.
Mazerolle, M. and M-A. Villard. 1999. Patch characteristics and landscape context as predators of species presence and abundance: a review. Ecoscience 6(1) 117-124.
McArthur, R. and E.O. Wilson. 1967. The theory of island biogeography. Princeton: Princeton University.
McGarigal, K. and W. McComb. 1995. Relationships between landscape structure and breeding birds in the Oregon coast range. Ecological Monographs 65(3): 235-260.
McGarigal, K. and B. Marks. 1995. FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. Portland (OR): USDA Forest Service, Pacific Northwest Research Station; General Technical Report PNW-GTR-351.
Merriam, G. 1988. Landscape dynamics in farmland. Trends in Ecology and Evolution 3: 16-20.
Miller, J., R. Brooks, M. Croonquist. 1997. Effects of landscape patterns on biotic communities. Landscape Ecology 12:137-153.
Mills, S. 1995. Edge effects and isolation: red-backed voles on forest remnants. Conservation Biology 9(2): 395-403.
Mladenoff, D. and H. He. 1999. Design, behavior and application of LANDIS, an object-oriented model for forest landscape disturbance and succession. In Spatial Modeling of Forest Landscape Change: Approaches and Applications. Mladenoff, D. and W. Baker (Eds.), Cambridge: Cambridge University Press.
Nikora, V., C. Pearson, and U. Shankar. 1999. Scaling properties in landscape patterns: New Zealand experience. Landscape Ecology 14(1): 17-33.
Nilsson, S., U. Arup, R. Baranowski, and S. Ekman. 1995. Tree-dependent lichens and beetles as indicators in conservation forests. Conservation Biology 9(5): 1208-1215.
Noss, R., and A. Cooperrider. 1994. Saving Natures Legacy: Protecting and Restoring Biodiversity. Washington: Island Press.
Opdam, P., R. Foppen, R. Reijnen, and A. Schotman. 1996. The landscape ecological approach in bird conservation: integrating the metapopulation concept into spatial planning. Ibis 137: 139-146.
Opdam, P. 1991. Metapopulation theory and habitat fragmentation: a review of holarctic breeding bird studies. Landscape Ecology 5(2): 93-106.
Ouborg, J. 1993. Isolation, population size and extinction: the classical and metapopulation approaches applied to vascular plants along the Dutch Rhine-system. Oikos 66: 298-308
Ortega, Y. and D. Capen. 1999. Effects of forest roads on habitat quality for ovenbirds in a forested landscape. The Auk 116(4): 937-946.
Paton, P. 1994. The effect of edge on avian nest success: How strong is the evidence? Conservation Biology 8(1): 17-26.
Plotnick, R. R. Gardner, and R. O'Neill. 1993. Lacunarity indices as measures of landscape texture. Landscape Ecology 8: 201-211.
Porneluzi, P., Bednarz, J., Goodrich, L., Zawada, N., and Hoover, J. 1993. Reproductive performance of territorial ovenbirds occupying forest fragments and a contiguous forest in Pennsylvania. Conservation Biology 7(3): 618-622.
Reunanen, P., M. M`nkkonen, and A. Nikula. 2000. Managing boreal forest landscapes for flying squirrels. Conservation Biology 14(1), 218-226.
Ripple, W., D. Johnson, K. Hershey, and E. Meslow. 1991. Old-growth and mature forests near spotted owl nests in western Oregon. Journal of Wildlife Management 55(2): 316-318.
Ritters, K., R. O'Neill, C. Hunsaker, J. Wickham, D. Yankee, S. Timmins, K. Jones and B. Jackson. 1995. A factor analysis of landscape pattern and structure metrics. Landscape Ecology 10(1): 23-39.
Robbins, C.S., D. Dawson, and B. Dowell. 1989. Habitat area requirments of breeding forest birds of the Middle Atlantic States. Wildlife Monographs 103: 1-34.
Robinson, S., R. Thompson, T. Donovan, D. Whitehead, D., and J. Faaborg. 1995. Regional forest fragmentation and the nesting success of migratory birds. Science 267: 1987-1990.
Roland, J. and P. Taylor. 1997. Insect parasitoid species respond to forest structure at different spatial scales. Nature 386: 710-713.
Roland, J. and P. Taylor. 1995. Herbivore-natural enemy interactions in fragmented and continuous forests. In Population Dynamics: New Approaches and Synthesis. Cappuccino, N. and Price, P. (Eds.), Academic Press, Pp. 195-208.
Roland, J. 1993. Large-scale forest fragmentation increases the duration of tent caterpillar outbreak. Oecologia 93: 25-30.
Saunders, D., R. Hobbs, and C. Margules. 1991. Biological consequences of ecosystem fragmentation: A review. Conservation Biology 5(1): 18-32.
Schmiegelow, F., C. Machtans, and S. Hannon. 1997. Are boreal birds resilient to forest fragmentation? An experimental study of short-term community responses. Ecology 78 (6): 1914-1932.
Schumaker, N. 1992. Using landscape indices to predict habitat connectivity. Ecology 77: 1210-1225.
Stanley, B., and P. Arp. 1998. Timber Harvesting of Forested Watershed: Impacts on Stream Discharge. Fredericton: Faculty of Forestry and Environmental Management.
Simberloff, D., and N. Gotelli. 1984. Effects of insularisation on plant species richness in the prairie-forest ecotone. Biological Conservation 29: 27-46.
Sorrell, J.P. 1998. Using Geographic Information Systems in the Cataraqui Watershed. Faculty of Environmental Studies, York University. http://www/nhb.com/sorrell/backgrd.htm
Storfer, A. and A. Sih. 1998. Gene flow and ineffective antipredator behavior in a stream-breeding salamander. Evolution 52(2): 558-565.
Tiebout, H., and R. Anderson. 1997. A comparison of corridors and intrinsic connectivity to promote dispersal in transient successional landscapes. Conservation Biology 11(3): 620-627.
Tinker, D., C. Resor, G. Beauvais, K. Kpfmueller, B. Fernades, and W. Baker. 1998. Watershed analysis of forest fragmentation by clearcuts and roads in a Wyoming forest. Landscape Ecology 13: 149-165.
Trzcinski, M., L. Fahrig, and G. Merriam. 1999. Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecological Applications 9(2): 586-593.
van Apeldoorn, R.C., C. Celada, and W. Nieuwenhuizen. 1994. Distribution and dynamics of the red squirrel (Sciurus vulgaris L.) in a landscape with fragmented habitat. Landscape Ecology 9(3): 227-235.
Verboom, J., A. Schotman, P. Opdam, and A..Metz. 1991. European nuthatch metapopulations in a fragmented agricultural landscape. Oikos 61: 149-156.
Villard, M., G. Merriam, and B. Maurer. 1995. Dynamics in subdivided populations of neotropical migratory birds in a fragmented temperate forest. Ecology 76(1): 27-40.
Waldick, R., 1994. Implications of Forestry-associated Habitat Conversion on Amphibians in the Vicinity of Fundy National Park, New Brunswick. Unpublished Masters Thesis Dalhousie University, Halifax.
Weinberg, H. and R. Roth. 1998. Forest area and habitat quality for nesting wood thrushes. The Auk 115(4): 879-889.
Winter, M. and J. Faaborg. 1999. Patterns of area sensitivity in grassland-nesting birds. Conservation Biology 13(6): 14241436.
With, K. and A. King. 1999. Dispersal success on fractal landscapes: a consequence of lacuniarity thresholds. Landscape Ecology 14: 73-82.
Whitcomb, B., R. Whitcomb, and D. Bystrak. 1977. Island biogeography and "habitat islands" of eastern forest III. Long-term turnover effects of selective logging on the avifauna of forest fragments. American Birds 31: 17-23.
Woodley, S. and G. Forbes. (Eds.). 1997. Forest Management Guidelines to Protect Native Biodiversity in the Fundy Model Forest. New Brunswick Cooperative Fish and Wildlife Research Unit, University of New Brunswick, 32 p.
Woodley, S., and G. Forbes. 1998. Forest management guidelines to protect native biodiversity. In the Fundy Model Forest. In State of the Greater Fundy Ecosystem. Woodley, S. Forbes, G., and Skibicki, A. (Eds.) Fredericton: Greater Fundy Ecosystem Research Group.
Woodley, S. 1993. Assessing and monitoring ecological integrity in parks and protected areas. University of Waterloo: Unpublished Ph.D. Thesis.
Zheng, D., D. Wallin, and Z. Hao. 1997. Rates and patterns of landscape change between 1972 and 1988 in the Changbai Mountain area of China and North Korea. Landscape Ecology 12: 241-254.
Click to Return to the Main Page